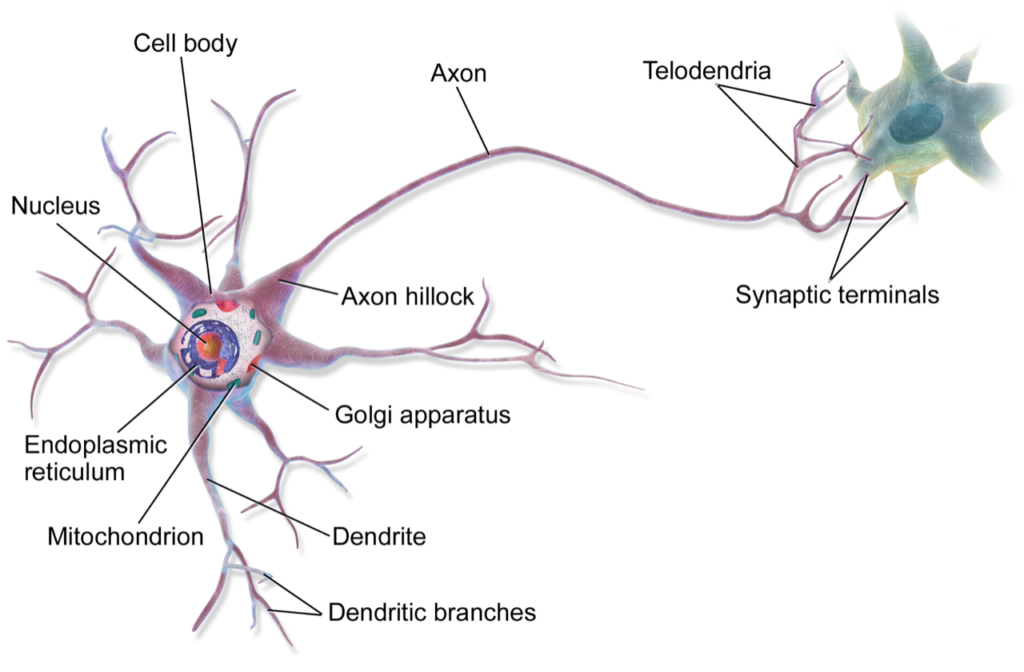
Fascia is the envelope that the message of consciousness arrives in. As we watch neurology advance to its next digitally-powered chapter, fascia and consciousness snuggle ever closer to each other.
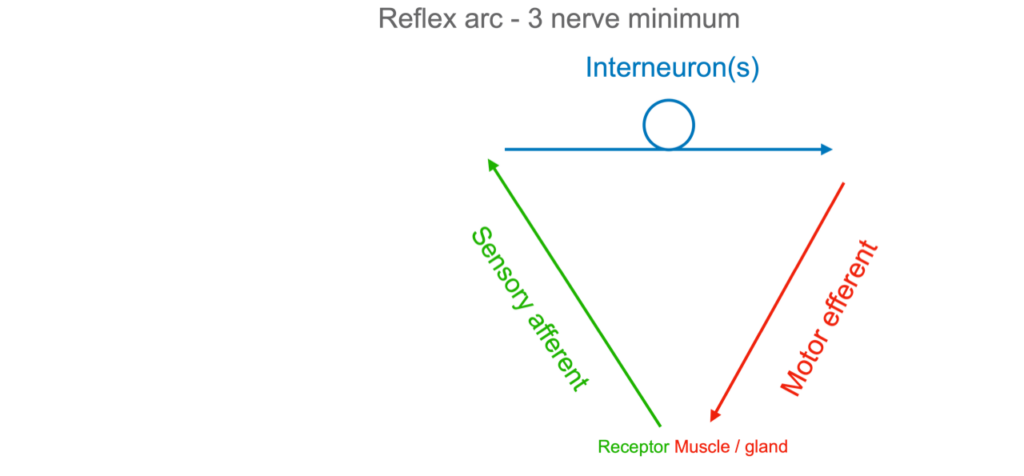
Neurology’s previous chapter – the one that I grew up on, exemplified in Skinner’s behaviourism (https://en.wikipedia.org/wiki/Behaviorism) – was based on the idea of ˆtabula raaa’ . With us since Aristotle, this idea is that that your baby’s mind is a ‘clean slate’. Onto the baby’s blank slate are projected incoming sensory signals (green), and out of which comes signals to muscles (red).
In between is always at least one interneuron (blue): When your GP strikes your infrapatellar tendon with a rubber hammer, the sensory signals from the quad muscle spindles run to the spinal cord, which is the first access to the central nervous system (CNS). With only one interneuron, the response is a reflex. By the time your brain knows about the sudden stretch, you’ve already extended your knee. You can dampen it ‘from above’, but you cannot stop it – with only one neuron in between, it is a physical reflex, not a conditioned one.
Most sensorimotor responses involve a great number of sensory inputs and an uncountable number of interneuronal loops (indicated by the blue circle – could be a few neurons, could be billions. In fact we all have friends, don’t we?, who seek our advice, only to have your good counsel get lost in the interneurons and never produce any salient motor output.
Please note the following:
• Sensory nerves provide data from both outside and inside the body. The teleceptors (eyes , ears, and nose) and somesthetic (skin) receptors bring us news from the outside. Many interoceptors lie in the gut, blood, lung, and kidney tubes, as well as all over your body to detect temperature and pain. Proprioceptors and the vestibular system (and vision when available) orient us in space.
• It is the interneuronal network, the CNS, that assesses all this data and initiates the motor output. The CNS interneurons have no actual contact with the outside world, only access to signals from end organs / receptors. They / It (what is your preferred pronoun for groups of neurons acting as one?) receive signals and send signals, abstractly constructing the world from these coded messages.
• Likewise, the CNS has no direct contact with the muscular movement itself – it relies on the change in sensory input to reflect and refine the movement intended when the signals were sent..
The contraction of the muscle (smooth, striated glandular, or cardiac) changes the sensation coming up the sensory nerves, and hence the triangle: sensation -> assessment -> motor response, sensation -> assessment -> motor response, sensation -> assessment -> motor response in endless loops that build to the complexity of skill shown on the athletic field or a climbing wall – or tying your shoes, a pretty complicated operation when you’re three.
So in the tabula rasa concept, everything starts wiped clean, in Alan Turing’s words, ‘much like the notebook one buys from the stationer’. In this model, as sensory input increases and gets integrated, the motor output becomes more organised and purposeful – and we call that ‘learning’. Input builds experience, experience builds competence and connection, et voila! a functioning adult emerges.
Preferred inputs put through the black box will beget preferred behaviours, and social evolution does the rest – so the theory goes. So hug your child, let them cry themselves to sleep – all these are inputs that are trying to produce the desired output, in other words, behave!
What went unnoticed – or got noticed and dismissed with a label – is that the brain, the CNS is left as a black box. Who decides what input results in what response? The church posits a ;’soul’, Descartes postulated a homunculus – the person within the person – and others have called it free will, executive function, intervening variables, or the decider. All of these terms are substitutes for the soul, the person, the mind, consciousness – fundamentally unscientific.
René Descartes – of ‘I think, therefor I am’ fame – postulated that the pineal gland (which can look for all the world like a ‘little man’ with arms and legs spread across the 4th ventricle. For Descartes, the homonculus runs our body like a puppet master. But who runs the puppet master.
The model, though, was simple and elegant and allowed behaviourists to learn many things about how we learn. But as we go to study consciousness, insubstantial and ungrounded concepts lie at the core of the tabula rasa and this whole neurological construct. It’s impossible to carry this argument any further, given the tenets that we grew up under.
Recognising the hole in the middle of our theory, scientific minds have been looking for alternatives. One that appeals posits an ‘inside-out’ theory of sense-making, rather than the ‘outside-in’ theories that fueled behaviourism, the ones we’ve been talking about until now.
I am very interested these days in how we create internal ’space’ in our minds. This refers to something very near our work – how we feel our body in gravity and how we negotiate acting within different environments. And yet there’s something just as elastic and just as important, but more subtle: How do we ‘construct’ the space we feel around us?
For example, imagine that you were reading your child a bedtime story. The house is quiet, the light low, the ‘space’ you create is very close and warm, a kind of cocoon all around you. Suddenly, there is a loud knocking on your front door. Immediately your sense of space elasticises, reaching down the stairs to make sense out of the sounds. The child’s room may feel suddenly cold and small – your sense of the space you’re in has changed dramatically. A trip down the stairs to discover your neighbour needs to borrow your car, and the world – internal and external – returns to its ‘proper’ size.
Similar but less dramatic elasticity of the perception of internal and external space accompany our day from the car to the office to the playing field. How our brains orient in space, the sense we make of the space around us – this realm bridges a gap from biomechanics to psychology. Many of the advances in performance and rehabilitation, in this new digital age of neurology, will be doctoring this sense of space and how we occupy it.
But we need to understand it first. Read the article referenced above, which accounts for this elasticity of perception, to get the ‘inside-out’ theory in some detail, but here’s my synopsis:
The signal process unit (CNS) starts out at birth with all of its machinery running. It is not waiting for experience to turn it on, it has tons of genetically created structure and sophisticated function online already. It is building new channels lickety-split on top of these inborn connections and those forged from sensations and chemistry in the months in the womb. Now it gets reams of sensory experience from the first contraction of labour, from the first breath forward.
In this process, there are two kinds of neurons, the ‘rich club’- the 20% that have many strong connections dispersed all over, who are disinclined toward plastic change – ergo very stable. 80% lie in the ‘poor club’, this lot lives with only a few local connections, and weak at that. The rich ones (these hard-working rich handle half the brain’s activity) maintain highly stable networks that ‘handle’ most situations. Novel events or sensations that cannot be quickly be resolved or ‘understood’ by these ‘tried and true’ solutions then involve the ‘poor club’ neurons, because they can plastically change their synapses to evolve new patterning – new ‘learning’. We’re not building a dictionary word by word, we arrive with a full dictionary full of nonsense, and it is slowly refined in this signaling world until it does make sense.
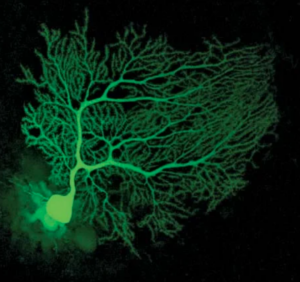
A Purkinje cell is an example of the ‘poor’ club, with many, many weak connections – and thus the ability to ‘learn’ – to make new connections.
Whereas this more ’standard’ neuron – 1 or two strong fixed connections – so: very reliable, not very ’tezchable’.
Importantly for our consciousness, the rich club set off waves of ‘corollary discharge’, which pre-inform other parts of the brain of what’s about to happen. Even as you read back and forth on these lines, neither the page nor your surroundings appear to move. Your brain was pre-informed of what was about to happen, and compensates. Close one eye and push the other eyeball from the side – does this page appear to move? Yes, it does. Because your finger isn’t set up to do these corollary discharges like the eye is,
This process of diverse and ‘holographic’ corollary discharge through the rich club neurons is essential for perception. You don’t ‘receive’ perception, you ‘do’ perception. You know how a still image can fool you for a minute – what is it? But small movement of your head resolves the anomaly and you ‘perceive’ what’s really there.
Boil it down to a phrase: Sensing + Doing = Meaning.
Sensing alone doesn’t make for perception, and in fact can often offer illusion Moving performed in the absence of sensation – all those folks with their earphones in jogging away on the hotel treadmills come to mind – can be meaningless. Only sensing coupled with movement – such as is being developed now by the spatial medicine experts, we trainers and manual therapists of all stripes – can produce meaningful experience.
 So in the tabula rasa concept, everything starts wiped clean, in Alan Turing’s words, ‘much like the notebook one buys from the stationer’. In this model, as sensory input increases and gets integrated, the motor output becomes more organised and purposeful – and we call that ‘learning’. Input builds experience, experience builds competence and connection, et voila! a functioning adult emerges.
So in the tabula rasa concept, everything starts wiped clean, in Alan Turing’s words, ‘much like the notebook one buys from the stationer’. In this model, as sensory input increases and gets integrated, the motor output becomes more organised and purposeful – and we call that ‘learning’. Input builds experience, experience builds competence and connection, et voila! a functioning adult emerges. Sensing alone doesn’t make for perception, and in fact can often offer illusion Moving performed in the absence of sensation – all those folks with their earphones in jogging away on the hotel treadmills come to mind – can be meaningless. Only sensing coupled with movement – such as is being developed now by the spatial medicine experts, we trainers and manual therapists of all stripes – can produce meaningful experience.
Sensing alone doesn’t make for perception, and in fact can often offer illusion Moving performed in the absence of sensation – all those folks with their earphones in jogging away on the hotel treadmills come to mind – can be meaningless. Only sensing coupled with movement – such as is being developed now by the spatial medicine experts, we trainers and manual therapists of all stripes – can produce meaningful experience.